Prokaryotik Kromozomlar: Prokaryotik Kromozomlar Üzerine Faydalı Notlar
Prokaryotik Kromozomlar hakkındaki notlarınız!
Prokaryotik bir hücrenin genetik bilgisi, bakteri kromozomu adı verilen yapı üzerindeki nükleolazmada taşınmaktadır. Asla bazik proteinlerle birleşik olmayan çift sarmal bir DNA (deoksiribonükleik asit) molekülünden oluşur ve bazı prokaryotlarda dairesel olduğu gösterilmiştir.

Resim Nezaket: hangthebankers.com/wp-content/chromosome.jpg
Sonuç olarak bakteriyel kromozom yapısal olarak ökaryotik hücrenin nükleer kromozomları ile değil, ökaryotik mitokondri ve kloroplastlarda bulunan organel DNA ile yapısal olarak homologtur. Tek bir bakteriyel kromozomun (yani, çok uzun bir DNA molekülü) prokaryotik hücrenin temel özelliklerini belirtmek için gerekli tüm genetik bilgiyi taşıması muhtemeldir.
Plazmidler: 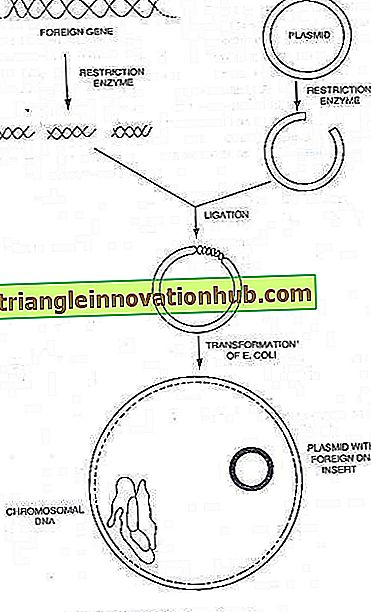

Pek çok bakteri ayrıca, bugüne kadar araştırılan plazmitler olarak bilinen, özerk replikasyon yeteneğine sahip, küçük, ekstra kromozomal dairesel DNA molekülleri barındırabilir, ilaçlara ve diğer antibakteriyel maddelere karşı direnç gibi bazı fenotipik karakterlerin ve bazı metabolik yollara aracılık eden enzimlerin belirleyicilerini taşır . Bir plazmiddeki DNA miktarı, bakteriyel kromozomdaki miktarın yüzde 0.1 ila 5'i kadardır.
Bu, Escherichia coli'de iki çiftleşme tipi olduğu ve konjugasyon sırasında bir eşin sadece genetik donör veya erkek, diğerinin sadece genetik alıcı veya dişi olarak davrandığı keşfedildi. Erkeğin tek işlevi, bazı DNA'sını aktarmak olduğundan, canlı kalması gerekmez, oysa dişi hücre, canlı kalmalıdır, böylece zigot içinde gelişebilir.
Bu, erkek ve dişi bakteriler konjuge olduğunda her dişi hücrenin bir erkeğe dönüştürüldüğü zaman bakterilerdeki erkekliğin bulaşıcı genetik bir element tarafından belirlendiği keşfedildi. Kalıtsal erkeklik özelliğini yöneten genetik element F faktörü (F = doğurganlık) olarak adlandırılır, sadece doğrudan hücreden hücreye temasla bulaşır. 1952'de Lederberg, plazmid terimini, F örneği olan tüm ekstra kromozomal kalıtsal belirleyiciler için genetik bir isim olarak kullandı.
Artık bakteri plazmitlerinin, genleri kendi replikasyonları için taşıyan küçük, dairesel DNA molekülleri olduğu bilinmektedir. Çoğu durumda, konakçı hücrede ilaçlara direnç veya toksinlerin üretilmesi gibi yeni özellikler kazandıran genleri de taşırlar. Pek çok plazmit, konjugasyon sürecini yöneten genleri taşır. Bu nedenle konjugasyon, bakteriyel hücreye bir plazmid tarafından uygulanan, normal sonucu plazmid DNA'nın transferi olan bir mekanizmadır.
Plazmid türleri:
Çoğu plazmit, konakçı özelliklerine göre sınıflandırılmıştır. Böylece, R faktörü (R = direnç) ve gram negatif bakterilerin COI faktörleri (COI = kolikinojen), Staphylococcus aureus'un penisilinaz plazmitleri, Pseudomonas'ın degradatif plazmitleri, kriptik plazmitleri vs.
Plazmidlerin özellikleri:
Bilinen tüm plazmidlerin yapısı, dairesel, çift sarmallı DNA moleküllerinden oluşur. Birçoğunun 5 x 106 ila 7 x 106 arasında moleküler ağırlıkları vardır. Bir (bir R faktörü) moleküler ağırlığa sadece 1 x 106 sahiptir ve kriptik plazmidlerin bazıları daha da küçüktür. 40, 000 moleküler ağırlığa sahip ortalama bir polipeptidi kodlamak için gereken DNA miktarı yaklaşık 6 x 106 olduğundan, F1 ve benzer büyüklükteki diğer plazmidler 100 kadar gen içerebilir.
Prokaryotlarda Gen İfadesi:
Bakteriyel kromozomlar:
Bakteriyel hücrenin merkezinde her zaman mitotik ve mayotik bölünme kabiliyetine sahip olmayan ve belirli bir nükleer zar içine alınmayan bakteriyel bir kromozomun bulunduğu her zaman tespit edilmiştir. Ortalama bir bakteri hücresi, tipik bir ökaryotik hücrenin DNA içeriğinin binde birini içerir.
Bakteriyel kromozom, az sayıda proteinle ilişkili ince uzun ve esnek bir DNA molekülü filamentidir. Bakteriyel hücreler çok hızlı bir şekilde bölünebilirler. Hücre bölünmesi, tüm hücre bileşenlerinin ikiye katlanması ve ardından hücrenin iki yeni hücreye bölünmesi ile tamamlanır. Hücre bölünmesindeki ilk adım DNA çoğaltmasıdır.
Bakterilerde Genetik Rekombinasyon:
Bu, iki ayrı genomda bulunan genetik materyallerin bir birim içerisinde bir araya getirildiği bir işlemdir. Bakterilerde rekombinasyon (1) transformasyon, (2) transdüksiyon ve (3) konjugasyon ile gerçekleşir. Bu fenomenler önceki paragraflarda, 'bakterilerde cinsel üreme' başlığı altında detaylı olarak tartışılmıştır.

İndüksiyon ve Baskı:
Bir virüsün genetik potansiyeli sınırlıdır ve esas olarak çoğaltma için ev sahibi makine kullanır. Öte yandan, bakteriler bileşenlerini basit tuzlardan ve şekerden sentezleyebilir. Bakteri Escherichia coli'nin kromozomunda yaklaşık 2500 gen bulunur. Bakteri sadece bir seferde birkaç genin hareketine izin vererek iç ekonomisini korur.
Bu, genlerin gereksinime göre açılıp kapatılacağı anlamına gelir. Yeni bir substratın işlenmesi ve metabolize edilmesi gerektiğinde bir dizi gen açılacaktır. Bu genlerin seti açıldığında, yeni substratı metabolize eden enzimler üretilir. Bu olguya indüksiyon denir ve bu olguyu uyandıran moleküller (metabolitler) indükleyicilerdir.
Öte yandan, bakteri tarafından ihtiyaç duyulan bir metabolit dışarıdan fazla miktarda sağlandığı zaman, bakteri bunu engeller ve bu da bakteri tarafından metabolitin daha fazla üretilmesini durdurur. Bu inaktive edilmiş genlere bu nedenle tekrar basılabilir olarak adlandırılır ve fenomen, baskı veya geri besleme baskısı olarak bilinir.
Bununla birlikte, bazı genler glikoliz gibi normal hücresel aktiviteye dikkat etmek için sürekli olarak eksprese edilir. Bu genler kurucu olarak bilinir ve bunlar tarafından üretilen enzimler dehidrojenazlar gibi kurucu enzimler olarak bilinir.
Operon Kavramı:
Francois Jacob ve Jacques Monod (1961), Paris Pasteur Enstitüsünde çalışırken, metabolik yolların bir birim olarak düzenlendiğini öne sürdü. Farklı tür şekerlerle donatıldıklarında, E. coli enzimlerinin niteliğindeki değişimi incelediler. Şeker laktozunun E. coli kültürlerine eklendiğinde, laktozun glikoz ve galaktoza parçalanması için gereken üç enzimi indüklediğini bulmuşlardır.
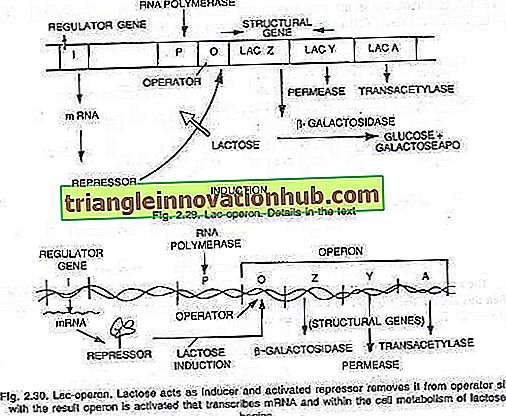
Bu yeni sentezlenmiş enzimler- (i) p-galaktosidaz, (ii) permeaz ve (iii) işlem-terazdır. Onlara göre, bu üç enzimin sentezi, bir operatör bölgesi O ve üç yapısal gen Z, Y ve A'ya bölünebilen operon olarak bilinen uzun bir DNA segmenti tarafından kontrol edilir. Yapısal genlerin etkisi, operatör bölgesi tarafından düzenlenir. Regülatör gen olarak bilinen 'i' geninin etkisiyle üretilen bir baskılayıcı proteinin yardımı.
Genler, operatör anahtarının açık veya kapalı olmasına bağlı olarak ifade edilir veya ifade edilmez. Anahtar açıkken, üç gen, RNA polimerazı ile, üç genin tümünü kapsayan tek bir mesajcı RNA dizisine kopyalanır. Her yapısal gen, genellikle cistron olarak bilinir ve tüm cistronları kapsayan kopyalanan uzun mRNA, polikistronik olarak bilinir.
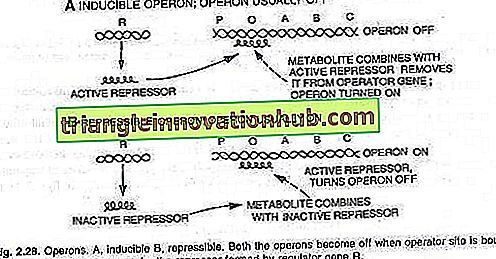
Operatör anahtarının açılması veya kapatılması, baskılayıcı olarak bilinen bir protein ile gerçekleştirilir. Bu protein operatöre (O) bağlandığında ve onu bloke ettiğinde, anahtar kapanır ve üç gen (Z, Y, A) eksprese edilmez, (bakınız şekil 2.28).
Lac-operon:
Bu uyarılabilir operonun bir örneğidir. Enzim geçirgenliğinin etkisiyle hücreye az miktarda laktoz molekülü eklenir ve az miktarda bastırılmış koşullarda bile bulunur. Bu birkaç molekül daha sonra, baskılayıcıya (i gen ürünü) bağlanan aktif bir laktoz formuna dönüştürülür ve bu nedenle baskılayıcı, operatör anahtarına daha fazla bağlanamaz.
Operatör, baskılayıcı RNA'dan uzak olduğunda polimeraz, promotor bölgesini P bağlayarak operonun transkripsiyonunu başlattığında, üç enzimin (P-galaktosidaz, permeaz ve işlemetilaz) üretilmesi için çevrilen üç enzime karşılık gelen Messenger RNA sentezlenir.
Bu üç enzimin üretimi ile laktoz metabolizması başlar. Enzimlerin sentezi, tüm laktoz moleküllerinin hücre tarafından tüketilmediği sürece ve sürece kadar devam eder. Baskılayıcıya bağlanan son laktoz molekülleri de tüketildiğinde, aktif olmayan baskılayıcı aktif hale gelir ve operonu normal şekilde kapatmak için operatör bölgesine (O) bağlanır (bakınız, şekil 2.29 ve 2.30).
Triptofan-operon:
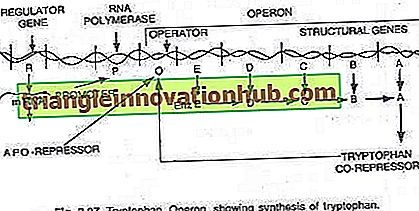
Bu durumda, operon genellikle açıktır, böylece transkripsiyon ve translasyon, hücre tarafından belirli bir metabolitin sentezi için gerekli enzimleri sentezlemek üzere açıktır. Bununla birlikte, hücre metabolit gerektirmediğinde veya metabolit fazla miktarda üretildiğinde, operon kapatılabilir. Triptofan operonu, triptofan (bir amino asit) sentezini katalize eden ve böylece oluşturan beş enzimi kodlayan beş genden (trp E, D, C, B ve A) oluşur bir anabolik yol. Triptofanın varlığı, üretiminden sorumlu enzimlerin sentezini bastırmaya hizmet eder (bakınız şekil 2.27 ve 2.31).
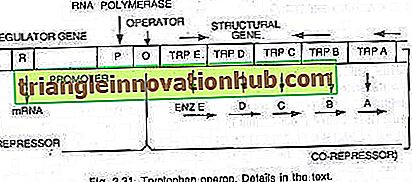
Bu beş enzimi kodlayan beş yapısal gen, tek bir operatör anahtarı tarafından düzenlenir. Burada, düzenleyici genler tarafından üretilen baskılayıcı protein, operatöre kendisi tarafından bağlanmaz. Bununla birlikte, triptofan eklendiğinde, baskılayıcıya bağlanır ve yardımcı baskılayıcı olarak bilinir. Repressor-corepressor kompleksi operatöre bağlanır ve bu operonun ekspresyonunu bloke eder.
Triptofan operonunun yapısı, lac operonunkine çok az ya da çok benzer, ancak fonksiyonel bir değişkenliğe sahiptir. Bu durumda, R gen ürünü, lak operonun i genine eşdeğerdir. R geni ürünü kendi başına operatöre bağlanamayan protein üretir. Buna apo-baskılayıcı denir.
Triptofanın bir eş-bastırıcısı varlığında, artık operatöre bağlanan ve operonun transkripsiyonunu ve triptofan üretimini önleyen fonksiyonel bastırıcı oluşturulur. Burada operon modeli prokaryotlarda hem indüksiyon hem de baskı fenomenini açıklar.